

ครูไวยุ์

ATP ถูกสร้างมาจากไหน??
กระบวนการสร้าง ATP อาจจะเกิดจาก
- Substrate level phosphorylation เป็นการสร้าง ATP จากสารพลังงานสูงอื่นๆ
- oxidative phosphorylation เป็นการสร้าง ATP จากสารนำอิเล็กตรอน (NADH และ FADH2) โดย NADH และ FADH2 สามารถถ่ายอิเล็กตรอนให้แก่ออกซิเจน โดยผ่านลูกโซ่ขนส่งอิเล็กตรอน (electron transport chain) ในไมโทคอนเดรีย (mitochondria) ทำให้เกิดพลังงานที่จะทำให้เกิดปฏิกิริยาระหว่าง ADP กับฟอสเฟตเป็น ATP
- photophosphorylation เป็นการสร้าง ATP จากพลังงานแสงสำหรับสิ่งมีชีวิตที่สังเคราะห์แสงได้ โดยพลังงานแสงจะทำให้น้ำแตกตัวให้ออกซิเจน โปรตอน และอิเล็กตรอน และอิเล็กตรอนจะถูกขนส่งไปตามลูกโซ่ขนส่งอิเล็กตรอนของคลอโรพลาสต์ทำให้เกิดพลังงานที่จะทำให้เกิดปฏิกิริยาระหว่าง ADP กับฟอสเฟตเป็น ATP
การเผาผลาญกลูโคสในอากาศให้พลังงาน 686 กิโลแคลอริ ต่อกลูโคสหนึ่งโมล (ประมาณ 180 กรัม) อุณหภูมิของร่างกายไม่สูงพอที่จะทำให้เกิดการเผาผลาญได้รวดเร็ว และปล่อยพลังงานออกมาอย่างมหาศาลในรูปของความร้อน แต่ในการออกซิไดส์กลูโคสในกระบวนการหายใจ ปฏิกิริยาไม่ได้เกิดขึ้นในขั้นตอนเดียวนั่นคือ ไฮโดรเจนพร้อม
อิเล็กตรอน จะไม่ถูกส่งต่อให้กับออกซิเจนไปพร้อมกันทั้งหมดในคราวเดียวแต่เกิดขึ้นเป็นขั้นเป็นตอนโดยอาศัยการทำงานของเอนไซม์ที่จะไปช่วยลดพลังงานกระตุ้นทำให้น้ำตาลกลูโคสถูกออกซิไดส์อย่างช้าๆ ที่อุณหภูมิร่างกายในกระบวนการหายใจ มีการขนย้ายอิเล็กตรอนเกิดขึ้นระหว่างการเกิดปฏิกิริยาเคมี การเปลี่ยนตำแหน่งของอิเล็กตรอน
ปลดปล่อยพลังงานที่สะสมอยู่ในโมเลกุลของอาหารออกมา พลังงานที่เกิดขึ้นนี่เองผลักดันให้เกิดการสร้าง ATP ขึ้น
การขนย้ายอิเล็กตรอนที่เกิดขึ้นระหว่างการเกิดปฏิกิริยาเคมีในกระบวนการหายใจระดับเซลล์ เราเรียกว่า ปฏิกิริยารีดอกซ์ (redox reaction) ในปฏิกิริยารีดอกซ์ การสูญเสียอิเล็กตรอนของโมเลกุลหรืออะตอมเรียกว่า การเกิดออกซิเดชัน (oxidation) ของโมเลกุลหรืออะตอมนั้น และการเติมอิเล็กตรอนให้กับโมเลกุลหรืออะตอมใดๆ ก็ตามเราเรียก การเกิดรีดักชัน (reduction) ของโมเลกุลหรืออะตอมนั้น

NAD+ เป็นตัวรับอิเล็กตรอนในบางช่วงของการหายใจ




(Electron transport chain)

1. NADH ส่งอิเล็กตรอนให้ complex I

สรุปปฏิกิริยาที่เกิดขึ้นในขั้นตอนนี้ คือ


การเผาผลาญกลูโคสในอากาศให้พลังงาน 686 กิโลแคลอริ ต่อกลูโคสหนึ่งโมล (ประมาณ 180 กรัม) อุณหภูมิของร่างกายไม่สูงพอที่จะทำให้เกิดการเผาผลาญได้รวดเร็ว และปล่อยพลังงานออกมาอย่างมหาศาลในรูปของความร้อน แต่ในการออกซิไดส์กลูโคสในกระบวนการหายใจ ปฏิกิริยาไม่ได้เกิดขึ้นในขั้นตอนเดียวนั่นคือ ไฮโดรเจนพร้อม
อิเล็กตรอน จะไม่ถูกส่งต่อให้กับออกซิเจนไปพร้อมกันทั้งหมดในคราวเดียวแต่เกิดขึ้นเป็นขั้นเป็นตอนโดยอาศัยการทำงานของเอนไซม์ที่จะไปช่วยลดพลังงานกระตุ้นทำให้น้ำตาลกลูโคสถูกออกซิไดส์อย่างช้าๆ ที่อุณหภูมิร่างกายในกระบวนการหายใจ มีการขนย้ายอิเล็กตรอนเกิดขึ้นระหว่างการเกิดปฏิกิริยาเคมี การเปลี่ยนตำแหน่งของอิเล็กตรอน
ปลดปล่อยพลังงานที่สะสมอยู่ในโมเลกุลของอาหารออกมา พลังงานที่เกิดขึ้นนี่เองผลักดันให้เกิดการสร้าง ATP ขึ้น
การขนย้ายอิเล็กตรอนที่เกิดขึ้นระหว่างการเกิดปฏิกิริยาเคมีในกระบวนการหายใจระดับเซลล์ เราเรียกว่า ปฏิกิริยารีดอกซ์ (redox reaction) ในปฏิกิริยารีดอกซ์ การสูญเสียอิเล็กตรอนของโมเลกุลหรืออะตอมเรียกว่า การเกิดออกซิเดชัน (oxidation) ของโมเลกุลหรืออะตอมนั้น และการเติมอิเล็กตรอนให้กับโมเลกุลหรืออะตอมใดๆ ก็ตามเราเรียก การเกิดรีดักชัน (reduction) ของโมเลกุลหรืออะตอมนั้น

NAD+ เป็นตัวรับอิเล็กตรอนในบางช่วงของการหายใจ
ในกระบวนการหายใจระดับเซลล์
เอนไซม์ในกลุ่มดีไฮโดรจีเนส (dehydrogenase) จะทำหน้าที่ในการถ่ายทอดอิเล็กตรอน จากน้ำตาลหรือสารอื่นๆ
ที่เป็นเชื้อเพลิงในเซลล์ไปยังตัวรับอิเล็กตรอน ซึ่งจะเก็บสะสมพลังงานเอาไว้กับตัว
ตัวรับอิเล็กตรอนในกระบวนการหายใจที่ชื่อ NAD+ ทำหน้าที่รับอิเล็กตรอนในหลายขั้นตอนของปฏิกิริยาการสลายกลูโคส
การรับโปรตอน 1 ตัวที่มาพร้อมกับอิเล็กตรอน 2 ตัวทำให้ NAD+ ถูกรีดิวซ์เป็น NADH ซึ่งจะผลักดันให้เกิดการสร้าง ATP เมื่อมีการถ่ายทอดอิเล็กตรอนจาก NADH ไปยังออกซิเจน
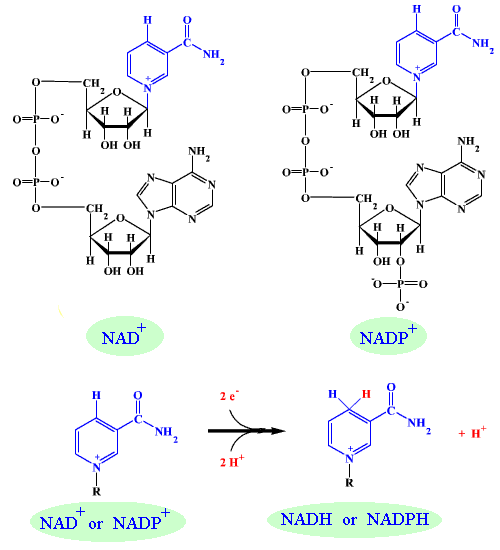
สารชีวเคมีที่สามารถรับและถ่ายอิเล็กตรอนได้ในเมแทบอลิซึม
Nicotinamide adenine dinucleotide หรือ NAD+ เมื่อรับอิเล็กตรอนแล้วจะอยู่ในสภาพรีดิวซ์ (NADH) เมื่อถ่ายอิเล็กตรอนแล้ว ก็จะกลับไปอยู่ในสภาพออกซิไดซ์ (NAD+) ได้เหมือนเดิม
Nicotinamide adenine dinucleotide phosphate หรือ NADP+ เมื่อรับอิเล็กตรอนแล้วจะอยู่ในสภาพรีดิวซ์ (NADPH) เมื่อถ่ายอิเล็กตรอนแล้ว ก็จะกลับไปอยู่ในสภาพออกซิไดซ์ (NADP+) ได้เหมือนเดิม
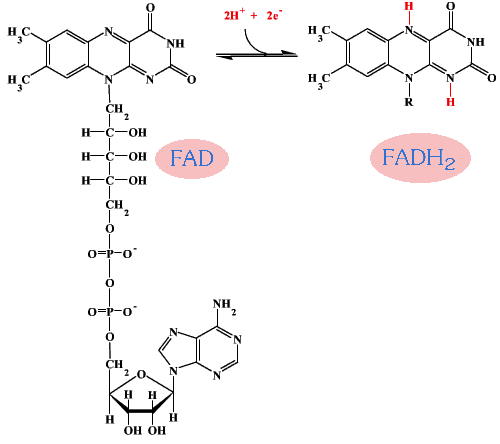
Flavin adenine dinucleotide หรือ FAD เมื่อรับอิเล็กตรอนแล้วจะอยู่ในสภาพรีดิวซ์ (FADH2) เมื่อถ่ายอิเล็กตรอนแล้ว ก็จะกลับไปอยู่ในสภาพออกซิไดซ์ (FAD) ได้เหมือนเดิม

กระบวนการสร้าง ATP จากน้ำตาลกลูโคส

(Electron transport chain)
เยื่อผนังชั้นในของไมโทคอนเดรียเชื่อมโยงการขนส่งอิเล็กตรอนเข้ากับการสร้าง ATP

Electron transport chain และ Chemiosmosis
องค์ประกอบของระบบถ่ายทอดอิเล็กตรอน คือ complex I, II, III และ IV ฝังตัวอยู่บนเยื่อหุ้มชั้นในของไมโทคอนเดรีย นอกจากกลุ่มโปรตีน 4 กลุ่มนี้ ยังมีโคเอนไซม์ Q และ ไซโตโครม c (cytochrome c) ซึ่งสามารถเคลื่อนที่ได้เพื่อช่วยในการถ่ายทอดอิเล็กตรอนระหว่างกลุ่มโปรตีน
กลไกการถ่ายทอดอิเล็กตรอนอาจแบ่งได้เป็น 4 ช่วง
กลไกการถ่ายทอดอิเล็กตรอนอาจแบ่งได้เป็น 4 ช่วง
1. NADH ส่งอิเล็กตรอนให้ complex I

การถ่ายทอดอิเล็กตรอนจาก NADH ให้โคเอนไซม์ Q ผ่าน complex I
ปฏิกิริยาใน complex I เกิดเป็นขั้นตอนดังนี้
1. NADH จะเข้าไปเกาะกับ complex
I และส่งอิเล็กตรอน 2 ตัว ให้แก่ FMN ทำให้ FMN ถูกรีดิวซ์กลายเป็น FMNH2
2. FMNH2 ให้อิเล็กตรอนต่อแก่ Fe-S ซึ่งเป็นกลุ่มของเหล็กและซัลเฟอร์หลายตัว ซึ่งสถานะของ Fe ใน Fe-S จะเป็น
Fe2+ เมื่อถูกรีดิวซ์ (รับอิเล็กตรอน) และเป็น Fe3+ เมื่อถูกออกซิไดซ์ (ให้อิเล็กตรอน)
3. Fe-S จะให้อิเล็กตรอนแก่โคเอนไซม์ Q
(Q ถูกรีดิวซ์กลายเป็น QH 2) การไหลของอิเล็กตรอน 2 ตัว
จาก NADH ไป โคเอนไซม์ Q
ทำให้เกิดปั๊มของโปรตอน ( H+) 4 ตัว
จากด้านในคือ แมทริกซ์ผ่านเยื่อหุ้มชั้นในออกไปที่ส่วนของช่องว่างระหว่างเยื่อหุ้มชั้นในและชั้นนอกของไมโทคอนเดรีย
สรุปปฏิกิริยาที่เกิดขึ้นในขั้นตอนนี้ คือ
NADH + Q + 5 H+matrix ---> NAD+ + QH2 +
4 H+cytosol
Q
หมายถึง โคเอนไซม์ Q
2. FADH2 ส่งอิเล็กตรอนให้ complex II

การถ่ายทอดอิเล็กตรอนจาก FADH2 ให้โคเอนไซม์ Q ผ่าน complex II
FADH2 ถูกสร้างขึ้นในวัฏจักรเครบส์จากการออกซิไดซ์ succinate ให้เป็น fumarate โดย succinate dehydrogenase ซึ่งเอนไซม์ตัวนี้เป็นส่วนหนึ่งของ succinate-Q-reductase หรือ complex II ในการนี้ FADH2 ที่เกิดขึ้นจะไม่หลุดออกจากเอนไซม์ ซึ่งเป็นส่วนหนึ่งของ complex II และจะส่งอิเล็กตรอน 2 ตัว จากโมเลกุลไปให้กลุ่ม Fe-S โดยตรง ซึ่ง Fe-S ก็จะส่งต่อให้โคเอนไซม์ Q (ทำนองเดียวกันกับที่เกิดขึ้นใน complex I) แต่ complex II จะต่างจาก complex I ตรงที่ไม่มีการปั๊มโปรตอน ( H+) ผ่านเยื่อหุ้ม ทั้งนี้เพราะพลังงานอิสระที่เกิดขึ้นจากปฏิกิริยาจะน้อยกว่าที่จะทำได้ ดังนั้น ปริมาณ ATP ที่จะสร้างขึ้นได้จากการออกซิไดซ์ FADH2 จะน้อยกว่าของ NADH
3. อิเล็กตรอนถูกถ่ายทอดจากโคเอนไซม์ Q ผ่าน complex III ไปยังไซโตโครม c

การถ่ายทอดอิเล็กตรอนจากโคเอนไซม์ Q ให้ไซโตโครม c ผ่าน complex III
complex III คือ Q-cytochrome c oxidoreductase บางครั้งเรียกว่า cytochrome b c1 complex เพราะ ประกอบด้วยไซโตโครม 2 ชนิด คือ b และ c1 (ซึ่งมี heme ที่มี Fe) และยังมีกลุ่มโปรตีน Fe-S เกาะอยู่ด้วย หน้าที่ของ complex III หรือ cytochrome reductase คือจะรับอิเล็กตรอน (2 ตัว) จากโคเอนไซม์ Q (QH2) และส่งต่อไปยัง cytochrome c และในขณะเดียวกันก็ปั๊มโปรตอน ( H+) ผ่านเยื่อหุ้มออกไปด้านนอก ในการถ่ายทอดอิเล็กตรอนจากโคเอนไซม์ Q ไปยัง complex III นั้นโคเอนไซม์ Q จะถูกออกซิไดซ์และรีดิวซ์ (Q และ QH2) สลับกันเป็นวัฏจักรที่เรียกว่า Q cycle
สรุปปฏิกิริยารวมที่เกิดขึ้นคือ
QH2 + 2Cytox +2H+matrix  Q + 2Cyt cred + 4 H+cytosol
Q + 2Cyt cred + 4 H+cytosol
4. อิเล็กตรอนถูกส่งจากไซโตโครม c ไปยัง O2 ผ่าน complex IV

complex IV รับอิเล็กตรอนจากไซโตโครม c และถ่ายทอดไปยัง O 2
complex IV คือ ไซโตโครม ออกซิเดส (cytochrome oxidase complex) หรือ cytochrome a a3 ประกอบด้วย heme A 2 ชนิดคือ a และ a3 ซึ่งเป็นฮีมที่มีทองแดง (Cu) เป็นองค์ประกอบ complex IV จะรับอิเล็กตรอนมาจากไซโตโครม c และส่งต่อให้ O2 อิเล็กตรอน 4 ตัว จะถูกส่งไปยังโมเลกุลของ O2 เพื่อรีดิวซ์ O2 ให้เป็น H2O ปฏิกิริยานี้เกิดพลังงานอิสระในปริมาณมากสำหรับปั๊มโปรตอนจากด้านแมทริกซ์ผ่านเยื่อหุ้มชั้นในออกไปด้านที่ติดกับไซโทซอล ซึ่ง proton gradient ที่เกิดขึ้นนี้ถูกนำมาใช้ในการสร้าง ATP
ปฏิกิริยาที่เกิดขึ้นคือ Fe2+ ในไซโตโครม c (ในรูปรีดิวซ์) 2 โมเลกุลส่งอิเล็กตรอนให้ Cu ของไซโตโครม a ซึ่งจะส่งต่อให้ Cu ของไซโตโครม a3 ซึ่งจะเป็นตัวส่งอิเล็กตรอนต่อให้ O2
รวมปฏิกิริยาที่เกิดขึ้นในขั้นตอนนี้คือ
4 Cyt cred + 8H+matrix + O2 4 Cyt cox + 2H2O + 4H+cytosol

การถ่ายทอด 2 อิเล็กตรอนจาก NADH หรือ FADH2 ไปยังโมเลกุลของ O2ทำให้เกิดพลังงานอิสระเท่ากับ -53 และ -36 kcal /mol ตามลำดับ กระบวนการออกซิเดทีฟ ฟอสโฟริีเลชัน จะจับพลังงานนี้ไว้ในรูปของฟอสเฟตพลังงานสูงของ ATP
NADH +  O2 + H+
O2 + H+  H2O + NAD+
H2O + NAD+  Gํ = -53 kcal/mol
Gํ = -53 kcal/mol
การที่กระบวนการออกซิเดทีฟ ฟอสโฟริเลชัน จะเกิดขึ้นได้นั้นขึ้นอยู่กับปัจจัยที่สำคัญ 2 อย่างคือ
ประการแรก เยื่อหุ้มชั้นในของไมโทคอนเดรีย จะต้องมีลักษณะที่ยอมให้ H+ ผ่านจากภายนอกเข้ามาภายในแมทริกซ์ได้โดยกระบวนการที่ต้องควบคู่กับการสร้าง ATP เท่านั้น
ประการที่สอง ความเข้มข้นของโปรตอนด้านนอกเยื่อหุ้มชั้นในจะต้องสูงกว่าภายในแมทริกซ์มาก (คือ เกิด proton gradient หรือ proton-motive force)

การปั๊มของโปรตอนผ่านเยื่อหุ้มชั้นใน
จากแมทริกซ์ไปยังช่องว่างด้านนอกที่ทำให้เกิดความแตกต่างของปริมาณโปรตอนใน 2 ด้านของเยื่อหุ้มและการที่โปรตอนไหลกลับผ่านทางช่องจำเพาะของ ATP synthase ทำให้เกิดการสร้าง ATP
จากแมทริกซ์ไปยังช่องว่างด้านนอกที่ทำให้เกิดความแตกต่างของปริมาณโปรตอนใน 2 ด้านของเยื่อหุ้มและการที่โปรตอนไหลกลับผ่านทางช่องจำเพาะของ ATP synthase ทำให้เกิดการสร้าง ATP
โปรตอนจะไหลกลับเข้ามาในส่วนแมทริกซ์ ผ่านทางโปรตีนที่ฝังอยู่บนเยื่อหุ้มชั้นใน โปรตีนตัวนี้คือ ATP synthase ( หรือบางทีเรียกว่า complex V) ATP synthase เป็นเอนไซม์ ประกอบด้วยหน่วยย่อย หลายหน่วย กลุ่มหนึ่งของหน่วยย่อยทำหน้าที่เป็นช่องให้ H+ ผ่าน อีกกลุ่มหนึ่งทำหน้าที่จับกับ ADP และPi เพื่อสร้าง ATP
แรงขับเคลื่อนโปรตอนทำให้เกิดการสร้าง ATP
ระบบถ่ายทอดอิเล็กตรอน และการสร้าง ATP โดย ATP synthase นั้น
เป็นคนละระบบแต่เกิดควบคู่กัน
โดยมีแรงขับเคลื่อนโปรตอน (หรือ proton gradient) เป็นตัวเชื่อมโยง
โดยมีแรงขับเคลื่อนโปรตอน (หรือ proton gradient) เป็นตัวเชื่อมโยง
Peter
Mitchell ซึ่งเป็นนักวิทยาศาสตร์รางวัลโนเบลใน
ค.ศ.1961 ได้ตั้งสมมุติฐาน chemiosmotic เพื่ออธิบายการสร้าง ATP ในไมโทคอนเดรีย
โดยกระบวนการออกซิเดทีฟ ฟอสโฟริเลชัน
การส่งอิเล็กตรอนต่อเป็นทอดๆ
ในระบบถ่ายทอดอิเล็กตรอนให้เกิดการปั๊มของโปรตอนจากข้างในแมทริกซ์ผ่านเยื่อหุ้มชั้นในออกไปสู่ด้านไซโทซอลของเยื่อหุ้มชั้นใน
(H+ ถูกปั๊มผ่านเยื่อหุ้มขณะที่มีอิเล็กตรอนไหลไปตามระบบถ่ายทอดอิเล็กตรอน)
ส่งผลให้ความเข้มข้นของ H+ ใน 2
ด้านของเยื่อหุ้มต่างกัน
คือทางด้านแมทริกซ์จะต่ำ และด้านช่องว่างระหว่างเยื่อหุ้มจะสูงและเกิดความต่างศักย์ที่เยื่อหุ้ม
โดยทางด้านแมทริกซ์จะเป็นลบ ทางด้านช่องว่างระหว่างเยื่อหุ้มเป็นบวก สมมุติฐานนี้เรียกว่า chemiosmotic hypothesis (ปฏิกิริยาเคมีเกิดควบคู่กับ osmotic
gradient) กล่าวคือ แรงขับเคลื่อนโปรตอน (proton-motive force) จะทำให้เกิดการสร้าง
ATP โดยเอนไซม์ ATP synthase ในสมมุติฐานนี้ การออกซิเดชัน และ
ฟอสโฟริีเลชัน (เติมหมู่ฟอสเฟต) เกิดขึ้นพร้อมกัน โดยมีแรงขับเคลื่อนโปรตอนเป็นตัวเชื่อมโยง
แรงขับเคลื่อนโปรตอนนี้เกิดขึ้นเนื่องจาก 2
ปัจจัย คือเกิดเนื่องจากความต่างศักย์ที่เยื่อหุ้มชั้นใน (V)
และเนื่องจากความแตกต่างของโปรตอนใน 2
ด้านของเยื่อหุ้ม (pH) ในกรณีของไมโทคอนเดรีย แรงขับเคลื่อนโปรตอนจาก V จะมากกว่าจาก pH
V)
และเนื่องจากความแตกต่างของโปรตอนใน 2
ด้านของเยื่อหุ้ม (pH) ในกรณีของไมโทคอนเดรีย แรงขับเคลื่อนโปรตอนจาก V จะมากกว่าจาก pH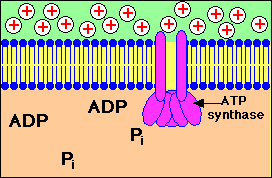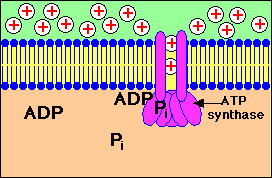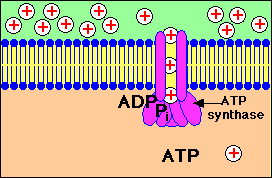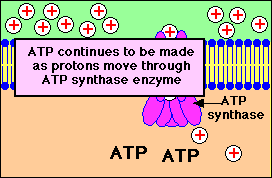
การคำนวณปริมาณ ATP ที่ได้กระบวนการถ่ายทอดอิเล็กตรอนและออกซิเดทีฟ
ฟอสโฟริเลชัน
1. ถ้าคิดจากสัดส่วน P:O เป็น 3 และ 2 สำหรับ NADH และ FADH2 ตามแบบเดิมในตำราทั่วไป การถ่ายทอดอิเล็กตรอนจาก NADH จะทำให้เกิด ATP 3 โมเลกุล และจาก FADH2 จะทำให้เกิด ATP 2 โมเลกุล ดังนั้นกระบวนการออกซิเดทีฟ ฟอสโฟริเลชัน จะให้ ATP 34 ตัว ฉะนั้น 1 โมเลกุลของกลูโคสจึงให้ 38 ATP
2. ถ้าคิดจากสัดส่วน P :
O ตามข้อมูลจากการวิจัยที่ทันสมัยขึ้น พลังงานจาก NADH
จะสามารถสร้าง ATP ได้ 2.5 โมเลกุล พลังงานจาก FADH2 จะสามารถสร้าง ATP ได้ 1.5 โมเลกุล
ดังนั้น ออกซิเดทีฟ ฟอสโฟริเลชันจะให้ ATP
28 โมเลกุล ฉะนั้น 1 โมเลกุลของกลูโคสจึงให้ 32 ATP
นอกจากการคำนวณแบบดังกล่าวนี้
ในปัจจุบันนี้ บางตำราคำนวณว่า กระบวนการออกซิเดทีฟ ฟอสโฟริเลชันให้ 26
ATP ฉะนั้น 1 โมเลกุลของกลูโคสจึงให้ 30 ATP
อย่างไรก็ตาม
การออกซิไดซ์กลูโคสอย่างสมบูรณ์นั้นจะได้พลังงานในรูป ATP โดยตรง เป็นส่วนน้อยคือ 4 ATP พลังงานส่วนใหญ่จะอยู่ในตัว NADH
(และ FADH2 เป็นส่วนน้อย) ซึ่งจะถูกนำไปใช้การสร้าง ATP ในกระบวนการออกซิเดทีฟ ฟอสโฟริเลชัน ดังที่ได้กล่าวมาแล้ว
อนึ่ง ปริมาณ ATP ที่ได้จากการออกซิเดชันของกลูโคสอย่างสมบูรณ์ตามที่กล่าวไว้ในตำราต่างๆ
นั้น มีตั้งแต่่ 30, 32, 36 และ 38
ATP ทั้งนี้ขึ้นอยู่ข้อมูลที่นำมาใช้ในการคำนวณ ตำราสมัยใหม่ในช่วง พ.ศ. 2546-47 นี้มักจะใช้ค่า 30-32 ATP มากกว่า 36-38
ATP แบบเดิม
ที่มา :
ไม่มีความคิดเห็น:
แสดงความคิดเห็น